Floración del maíz
En
el maíz los órganos florales femeninos y masculinos se ubican dos estructuras de la misma planta, la espiga y el jilote. Cada
espiga produce de 2 hasta 5 millones de granos de polen, esto significa entre
2,000 y 5,000 granos de polen disponibles por cada óvulo o su estigma
respectivo, están disponible después que el rocío se ha secado. Un día típico, el
polen se disemina entre las 9:00 y las 11:00 AM, cuando la temperatura es de 24.3
± 0.8 °C (Figura 1, Figura 2).
La espiga puede estar totalmente emergida y haber alcanzado su tamaño
final antes de que cualquier grano de polen haya sido diseminado, no siempre es
así. El polen es ligero y es transportado por el viento pero la mayor parte
de él se queda depositado en distancias que oscilan entre los 8 y 20 metros,
aunque se ha encontrado a más de 500 metros. Hay pocas posibilidades de que una lluvia deslave al polen depositado en los estigmas, estas están recubiertas
de un vello muy fino y pegajoso que les sirve para atrapar y anclar a los
granos de polen. El grano de polen permanecen viables sólo de 18 a 24 horas. Un
jilote bien desarrollado puede producir 1,000 óvulos (potenciales granos o
semillas de maíz).
Polinización
El polen derramado se deposita sobre los
estilos, que son receptivos a todo su largo, y por ser de carácter mucilaginoso
(húmedos y pegajosos), permiten la adherencia y su germinación. En cada
estilo germina varios granos de polen, pero sólo uno de ellos, penetrará al
interior a través de su tubo polínico; éste elonga hasta alcanzar la ovocélula y
fecundarlo.



La germinación de un grano de polen ocurre, a pocos minutos
después de que entra en contacto con el estilo, desde la
germinación del polen hasta el momento de la fecundación, transcurren entre
12 y 24 horas. La liberación de polen inicia
normalmente en las espiguillas de la mitad superior de la espiga central (Figura 3), y termina en el extremo apical de las espigas laterales más
bajas; en cada planta, la etapa de liberación de polen puede durar entre 7 y 10
días.

El máximo derramamiento de polen en una planta se produce 3 a 4 días después de ocurrida la antesis en las primeras anteras. En una mazorca, los primeros estilos que aparecen a través de las brácteas u hojas envolventes son los de la base del jilote; la aparición de los estilos en una planta comienza 1 a 2 días después que ha comenzado la liberación del polen. Cada mazorca requiere un promedio de 4 a 5 días para completar la emisión de sus estilos; éstos, a su vez, pueden lograr un crecimiento diario de 2.5 a 3.0 cm.

El máximo derramamiento de polen en una planta se produce 3 a 4 días después de ocurrida la antesis en las primeras anteras. En una mazorca, los primeros estilos que aparecen a través de las brácteas u hojas envolventes son los de la base del jilote; la aparición de los estilos en una planta comienza 1 a 2 días después que ha comenzado la liberación del polen. Cada mazorca requiere un promedio de 4 a 5 días para completar la emisión de sus estilos; éstos, a su vez, pueden lograr un crecimiento diario de 2.5 a 3.0 cm.
A pesar que la cantidad de polen
producido, pueden ocurrir fallas en la producción de granos,
especialmente en la parte apical de las mazorcas. Así, ante condiciones secas y
calurosas, o de déficit hídrico, los estilos pueden deshidratarse a tal nivel,
que su contenido de humedad se haga insuficiente para la germinación del polen. Las plantas no logran una suficiente homogeneidad, iniciando su
emisión de estilos en un período que normalmente se prolonga por 10 días o más.
En este sentido, las mazorcas que inician tardíamente su emisión de estilos
pueden verse afectada su polinización.
Granos de Polen
El gametofito femenino
El gametofito femenino o saco embrionario, se origina a partir de una gran célula diploide (2n) o célula madre de la megaspora. En la mayoría de las angiospermas está envuelta por una o dos láminas de tejido (tegumentos) que dejan un espacio abierto en el ápice denominado micropilo. Los tegumentos se unen en la zona basal, llamada calaza, por donde llegan los haces vasculares procedentes de la placenta, una zona muy vascularizada de la pared del ovario.
A partir de la célula madre de la megaspora (2n) se producen mediante una meiosis 4 megasporas (n) de las que sólo una será viable, esta megaspora es homologa al grano de polen recién formado con contenido aun unicelular. Aunque el modelo puede variar, en la mayoría de las angiospermas para formar el megagametofito de la megaspora (n) sufre tres mitosis sucesivas alcanzando un estado final de 8 núcleos (n) dispuestos en dos grupos de 4 que se sitúan cerca de los polos.
El paso final es donde uno de los núcleos de cada grupo, emigra al centro de la célula. Son los núcleos del endospermo que tendrán un papel relevante en la fabricación del tejido nutricio de la semilla. La célula central del grupo que está cerca del micropilo contiene a la ovocélula (gameto femenino) las otras dos se denominan sinérgidas y antípodas, ambas estériles.
La fecundación es un proceso que se inicia con el deposito o el contacto del grano de polen sobre el estilo de la flor, puede ser uno o varios granos, pero solo uno lo fecundará. El polen al caer sobre el estilo inicia la elongación del tubo polínico provocado por el núcleo vegetativo, durante su elongación del tubo polínico el núcleo generatriz presenta una mitosis formando dos células n, el tubo polínico se orienta por estímulos fitohormonales del megagametofito hacia el micrópilo, un núcleo generatriz se fusiona con la ovocélula el cual constituirá el embrión de la semilla (2n), y el segundo núcleo se fusionará con las dos células centrales, estás originarán una tejido triploide (3n) que posteriormente se transformarán en el endospermo de la semilla, este proceso se le denomina doble fecundación.
M.C. Francisco Chablé Moreno
Docente-Investigador del Instituto Tecnológico de Roque
Celaya, Guanajuato, México
Este documento se ha editado de imágenes de varios autores, se hace con la finalidad de realizar un entendimiento más fácil de la fecundación en las plantas angiospermas, pero se tratado de focalizar a la fecundación de maíz y la formación de los tejidos 2n y 3n.
Granos de Polen
La
disciplina que estudia el polen y las esporas se denomina palinología. El grano de polen posee dos núcleos, el vegetativo y el generativo. En el citoplasma del núcleo vegetativo del polen hay abundantes orgánulos y
reservas: RE (retículo endoplasmático), dictiosomas, plástidos con almidón que
se gasta en la formación del tubo polínico, lípidos, proteínas, vitaminas. Los
granos de polen forman tubos polínicos muy largos
generalmente tienen lípidos como sustancia de reserva; los polinizados
por animales tienen azúcares o aceites; los que son llevados por el viento
tienen frecuentemente almidón. La protección de su contenido está
asegurada por la presencia de una pared muy resistente, también llamada esporodermis. Esta pared contiene proteínas y enzimas, responsables de
las reacciones de incompatibilidad que ocurren entre el polen y el estigma. De
afuera hacia dentro se pueden distinguir la exina y la intina.
La intina envuelve
al protoplasma, es delicada, poco resistente, constituida de celulosa y
pectina, es más gruesa generalmente a la altura de las aperturas.
La exina está
constituida por esporopolenina, sustancia químicamente muy
resistente y solo degradable por oxidación. Es un polímero de carotenos y sus
ésteres. Estudios realizados en Liriodendron indican que los
precursores de la esporopolenina son sintetizados en el RE de las microsporas y
de las células del tapete (Gavarayeba, 1996).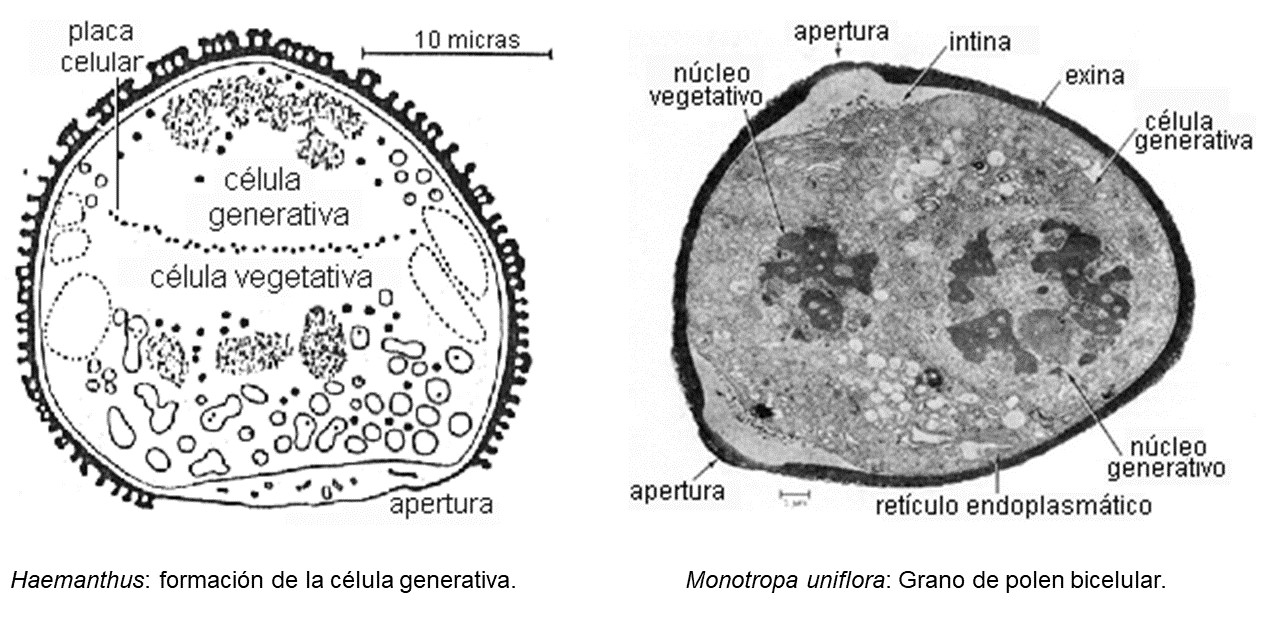 |
| Figura 4. El citoplasma de la célula generativa es menor en comparación a la célula vegetativa. |
El gametofito femenino
El gametofito femenino o saco embrionario, se origina a partir de una gran célula diploide (2n) o célula madre de la megaspora. En la mayoría de las angiospermas está envuelta por una o dos láminas de tejido (tegumentos) que dejan un espacio abierto en el ápice denominado micropilo. Los tegumentos se unen en la zona basal, llamada calaza, por donde llegan los haces vasculares procedentes de la placenta, una zona muy vascularizada de la pared del ovario.
A partir de la célula madre de la megaspora (2n) se producen mediante una meiosis 4 megasporas (n) de las que sólo una será viable, esta megaspora es homologa al grano de polen recién formado con contenido aun unicelular. Aunque el modelo puede variar, en la mayoría de las angiospermas para formar el megagametofito de la megaspora (n) sufre tres mitosis sucesivas alcanzando un estado final de 8 núcleos (n) dispuestos en dos grupos de 4 que se sitúan cerca de los polos.
El paso final es donde uno de los núcleos de cada grupo, emigra al centro de la célula. Son los núcleos del endospermo que tendrán un papel relevante en la fabricación del tejido nutricio de la semilla. La célula central del grupo que está cerca del micropilo contiene a la ovocélula (gameto femenino) las otras dos se denominan sinérgidas y antípodas, ambas estériles.
 |
| Figura 5. Diagrama esquematizado de la formación de la estructura del megagametofito. |
La fecundación es un proceso que se inicia con el deposito o el contacto del grano de polen sobre el estilo de la flor, puede ser uno o varios granos, pero solo uno lo fecundará. El polen al caer sobre el estilo inicia la elongación del tubo polínico provocado por el núcleo vegetativo, durante su elongación del tubo polínico el núcleo generatriz presenta una mitosis formando dos células n, el tubo polínico se orienta por estímulos fitohormonales del megagametofito hacia el micrópilo, un núcleo generatriz se fusiona con la ovocélula el cual constituirá el embrión de la semilla (2n), y el segundo núcleo se fusionará con las dos células centrales, estás originarán una tejido triploide (3n) que posteriormente se transformarán en el endospermo de la semilla, este proceso se le denomina doble fecundación.
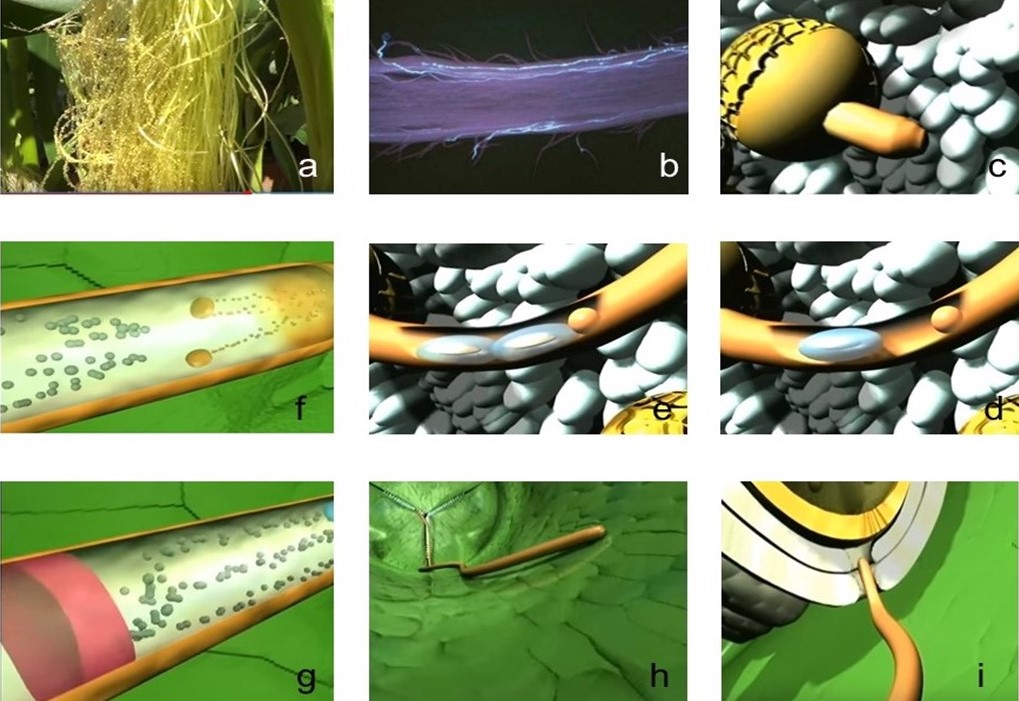 |
Figura 6. Proceso de elongación del tubo polínico e inicio de la fecundación vegetal.
|
M.C. Francisco Chablé Moreno
Docente-Investigador del Instituto Tecnológico de Roque
Celaya, Guanajuato, México
Este documento se ha editado de imágenes de varios autores, se hace con la finalidad de realizar un entendimiento más fácil de la fecundación en las plantas angiospermas, pero se tratado de focalizar a la fecundación de maíz y la formación de los tejidos 2n y 3n.
A disposición de mis estimados alumnos del Instituto Tecnológico de Roque
ResponderEliminarEste comentario ha sido eliminado por el autor.
ResponderEliminarEn que parte de la planta ocurre? o es en toda la planta?
ResponderEliminar